Rebirth of a Movement
By Will Brinton, Ph.D.
A renewed national interest in soil health and soil biology is creating an alternative to the long-held chemistry-based mineral theory of soil fertility still dominant today.
For about 175 years, soil has been viewed largely as a physical support medium for plants to which mineral nutrients must be supplied based on limiting factors and crop removal.
Along with this, soil testing labs have fine-tuned rapid soil chemical extractions as a means to recommend mineral supplements, based on a formidable foundation of mathematical equations calibrating to potential yields.
This “chemical-mathematical” approach has led to increased reliance on costly mineral fertilizer supplements, against a backdrop of declining nutrient efficiency connected to poorer economic returns and decreased soil health, a syndrome of compaction, salinization, decreased organic matter (OM) and loss of soil biology. A recent study in Nature by researchers at McGill University and University of Minnesota of trends of major crops across all major continents indicates yields are plateauing and in many cases now declining.
There is an interesting angle to mineral theory that comes closer to home in organic practices. This has to do with mineral balancing and using “cation exchange” theory as a means to amend soils. While organic growers reject one aspect of mineral theory, this one, which is entirely inorganic in origin, has gained a strong foothold. I view the practice of external balancing of minerals as essentially an import from conventional ag science – perhaps a necessary evil.
The alternative, emerging view sees soils as self-structuring systems in which biology regulates crop growth, releases and delivers nutrients and creates soil physical integrity – a niche in which microbes can flourish. This view requires new forms of “soil health” or soil biology tests, such as microbial CO2 respiration, water stable aggregates, earthworm counts, bacterial and fungal mass, water-soluble carbon and amino-nitrogen – not your ordinary set of lab protocols.
Focusing on soil biology should ultimately reduce fertilizer expenditures, improving nutrient efficiency by enabling plant and microbe relationships, reducing nitrate leaching to improve water quality, and reducing unnecessary soil tillage to aid in carbon sequestration and soil structure.
Farmers have increased use of soil-building “cover crops” (originally “green manures”) significantly – something many in organic farming implemented in the ‘70s and ‘80s. This soil-building system is becoming harder to grasp quantitatively, and it challenges soil labs, which may not have the models to account for it! Of many possible indicators, our lab has taken about five to focus on method development.
An interesting outcome of the growing awareness of soil building is recognition of the importance that including animals plays in farming systems due to synergistic benefits to soils, not to mention the huge significance to human diets of dramatically increased quality of meats from unstressed and healthier animals.
The following diagram juxtaposes the past-to-present and the newer, emergent views.
| PAST ABIOLOGICAL VIEW | EMERGING BIOLOGICAL VIEW |
| Soil is physical basis to support plants; mineral nutrients and minerals must be added according to limiting factors and crop removal demand | Soil functions as a self-structuring system; soil biology actively regulates crop growth and soil physical integrity |
| TOOLKITS SUPPORTING THE VIEW | |
| “SOIL CHEMICAL ANALYSIS” – Quick chemical extractions; mathematical calibration of tests to relative crop yield; balancing extracted soil mineral levels | “SOIL HEALTH AUDITS” – Soil respiration, soil earthworm counts, soil aggregate stability, bacterial and fungal mass, soil organic matter; soluble carbon, humus amino-N |
| RESULTS OF APPLYING THE APPROACH | |
| Increased reliance on costly nutrient inputs and “tweaking” minerals; lower net profit of yields; increased soil compaction; salinization; loss of organic matter; decrease in soil biology; unreliability of yields | Reduced reliance on fertilizers; more use of soil-building cover crops; improved N-efficiency, less P and N-leaching; increased carbon sequestration; more inclusion of animals; reduction in soil tillage; improved soil structure, improved yield reliability |
The Unseen World of Roots and Microbes
The role plant roots play in soil building is far more extensive and important than commonly thought. Root mass of plants closely mirrors above-ground plant mass. In a recent article titled “Moving beyond the soil test,” Bill Liebhardt, known for his early years at Rodale and more recently at UC Davis, writes that the roots of a single plant can explore up to 200 cubic feet of soil. However, soil test models assume that roots explore perhaps only about 10 cubic feet.
Many new efforts in root studies show that a single plant may have miles of roots when added end-to-end; calculated in three dimensions, the biological surface area can equal that of several baseball fields. If plants exert an influence on the soil, so does the soil exert one on plants. A Canadian study found that a single barley plant had 46 square feet of roots per cubic foot of poor soil, but 495 square feet in improved soil and 852 square feet in a healthy soil. From this it is easy to grasp that healthy roots “know” every nook and cranny of a soil, but under stressed conditions – for whatever cause – may be unable to access much of it.
The Road Not Taken: Soil Biology
Many people think there is one method of science, and that we have followed it consistently in agriculture into the present. Yet if we examine the history of science, we find evidence of another path in agricultural science not taken that could have been, in which case we might be in an entirely different situation presently.
In the late 1800s and early 1900s, interesting empirical results and some efforts to form new biological theories to improve yields can be found in pockets around the world, key examples being Sweden, Germany and Czechoslovakia. Notably, Swedish biologist Lundegårdh (1880-1955) performed innovative work observing soil biology functioning, traces of which are found in Waksman’s microbiological notes at Rutgers from the 1920s, but abandoned amid the stunning medical advances coming out his and Albert Shatz’s (my advisor’s) work in soil microbiology. Similarly, a scattering of advanced work in the United States and United Kingdom, also in the 1920s, advocated a scientific-biological soil management model, with soil metabolism as the focus. This differs sharply from what actually developed.
An example of an early effort to develop new soil tests to lead in another direction is the work on CO2 respiration by Isermeyer and Koepf at Hohenheim University (Germany) in the 1950s, leading to the first comparative studies of soil respiration in organic versus conventional systems. Unfortunately, those who promoted these early “soil health” methods were trampled in the excitement of fertilizer technology and resulting booming yields that resulted.
A startling example of excitement amid error is found in Justus von Liebig’s work from the 1860s, essentially launching the nutrient-fertilizer concept of plant nutrition – but without any notion of soil microbes. For all his success in chemistry analysis of plant ash, microbes and nitrogen fertility completely eluded him. He observed accurately that plants raised after legumes had more nitrogen content, but he was convinced that air was fertilizing plants with ammonia.
Famously, Liebig, in a debate with Pasteur, rejected the view that a fermenting mass could be “alive.” He presumed it and all plant nutrition to be an as-yet hidden chemical process. This is surprising today considering his stature and the innovative technological significance of many of his mineral and organic molecule discoveries. This error – and others like it – I believe virtually condemned the new field of plant nutrition to a firm, non-biological foundation.
Just when an inkling of a new biological science was possible early in the 20th century, Haber in Germany launched the chemical nitrogen-fixation era – a huge leap forward (intended principally to obviate the Allied Chilean nitrate blockade), and which was introduced wholesale to farming right after WWI (incidentally prompting some German farmers as early as 1920 to object, and this resulted in the birth of the Biodynamic farming movement). My view is that at this point science had already been consigned to a backseat in the face of exciting industrial developments thought at the time to be essential.
Meanwhile, Lundegårdh famously lamented in a published article in Soil Science in April 1926: “The effect of fertilizers increasing plant growth is the only one given any attention in agriculture.” The depth of meaning to this complaint is large when seen against the backdrop of his extensive work on soil biological functioning.
Early soil biological discoveries that may well have led to “organic farming before organic farming” suffered a large setback due to the one-sidedness of science leading into the war years, from which the inorganic-fertilizer industry and the science of “calibration amendments” emerged as a super-sized world industry. A second setback – minor in comparison – occurred in the mid-1970s when science supporting organic farming was growing, especially in Europe. To contest this was the newly formed Council for Agricultural Science and Technology (CAST), established in the United States by major land-grant universities, the USDA and the Agronomy Society. CAST had a stated mission to question the development of, and in my personal experience, restrict access to scientific information about, organic farming, founded on biological principles described a generation earlier. While unsuccessful in the end if not in fact energizing radicalism in organic farming, this union also set back the cause of biological science joining with agriculture.
I am concerned about lost time, since now so much more fertilizer, water and adjuvant chemical support is required to get the same yields in all major food crop cereals – a problem for the environment and for all soils of the world.
Soil Health and How Microbes Resist Measuring
A biologically-oriented soil model – the path not taken – offers an escape from the reductionist and downward spiraling industrial-science scenario characterized above.
By one accounting, soil organic matter can be broken down into three categories. The largest, comprising roughly 85 percent, is humus; 10 percent is plant roots in the process of decaying; and the balance of 5 percent may be living soil organisms. For over a century now, when soil labs have tested soil, we have combined all three in one “ashing” process reported as “OM” – a large simplification, since the biology of OM is important. The very modern preoccupation with “carbon” and carbon sequestration similarly overlooks the essential biology of this building block (while of course the new carbon focus may be essential in the global climate discussion).
Of the 5 percent of soil OM consisting of living soil organisms, perhaps 40 percent is estimated as algae and fungi, 40 percent bacteria and actinomycetes and 20 percent soil-inhabiting animals. That 20 percent is disrupted by tillage, while the other 80 percent is much less affected.
Most of the individual soil organisms taxonomically remain unknown because of the difficulty of isolating, culturing and identifying them; those that have been identified are primarily the medically important ones. We do know about overall numbers and mass, though. Hoorman and Islam from Ohio State University give one distribution of microbes per acre, which, converted to a mass basis, shows up to 7 tons of organisms per acre of farmland: in this case 36 pounds of nematodes, 45 pounds of protozoa, 134 pounds of algae, 1,780 pounds of fungi, 2,225 pounds of actinomycetes and 2,448 pounds of bacteria. A soil with 2.5 percent OM has a total mass of 50,000 pounds of pure OM per acre in the topsoil layer, feeding between 1 and 7 tons per acre of microbes – all respiring, breathing out CO2. In turn – and this reaches back to Lundergårdh’s work – this CO2 is associated with a large mass turnover of nutrients and the direct uptake for plant photosynthesis of carbon respired only moments before from the living soil.
The new Solvita test measures microbial respiration via a “CO2 burst” that comes from dried soils that have been rewetted. We developed it in our laboratory to provide a cost-effective method for soil testing labs to report this aspect of soil health. Incidentally, the established soil testing industry is another potential impediment to change, since it has so heavily invested in the mineral nutrition model from the past.
With Solvita we have worked to develop alternative tests to reveal other features, such as amino-N, a form of N derived from living organisms that is loosely sequestered in humus and yet is relatively available to plants given soil biological activity. Ordinary soil tests do not include any such estimate. Tests conducted at Woods End and University of Connecticut reveal anywhere from 300 to 500 pounds of amino-N/acre is present in soils where manure compost has been applied, correlating closely with plant yield and N-uptake where no other nitrogen fertilizer is used.
Nature’s Plan and Some Evidence
Earth’s green carpet is an extraordinary filter for grabbing CO2 coming directly out of the earth. In warm, moist conditions in a healthy, vibrant soil, microbes and plant roots release about 100 pounds of CO2 per acre per day. CO2 should be grasped as a major crop nutrient since it is required in large amounts for photosynthesis. Plants, it turns out – and this also goes back to Lundergårdh – are much more likely to get CO2 from the soil than from the atmosphere above them, especially if the soil is alive. How much CO2? Wheat, for example, absorbs up to 110 pounds of CO2 per acre per day; corn, up to 400 pounds, during full growth midsummer.
While atmospheric CO2 levels are now at 400 ppm, this is actually a very small amount from the perspective of growing plants. Beneath the ground inside the airspace of topsoil, the CO2 levels can reach 4,000 ppm, fluctuating with temperature and moisture; immediately above the soil surface in the air around lower leaves and vegetation, there might be about 1,500 down to 800 ppm, diminishing rapidly to ambient levels outside the actual canopy. Plants absorb this soil CO2, and the richer the soil, the more CO2 they may proportionally obtain from the soil as opposed to the atmosphere! A recent study in diverse-canopy tropical forests shows that none of the CO2 produced inside the forest ever gets out to the atmosphere – it’s all reabsorbed on the way up. This is the fundamental discovery as presented by Lundergårdh, and it is basically nature’s plan – clearly the solution to keeping atmospheric CO2 down is a green-covered earth! The focus on “soils will save us” should shift to “plants will save us,” and soil should never be bare.
When soils release CO2, nutrients that were associated with the carbon are also set in motion, including as much as 1 to 5 pounds of N per day in very warm, active soils. If sustained for even 30 days, that would be 50 to 150 pounds of N – perhaps enough to grow a crop. This is the essence of the organic view that the soil (microbes) feeds the plants.
| Figure 1 – Relation of Soil Biology to Yield in Absence of any Fertilizer |
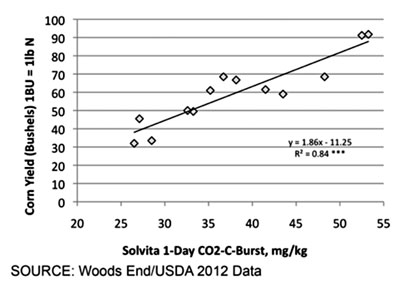 |
Quite a range of biology can be quantified when going from depleted to rich soils, even within a similar soil region, as shown in the following table from North Carolina. The results point to how past management may enhance or deplete soil biology. Results are contrasted with a virgin prairie soil from Nebraska:
| Table 1 – Soil Respiration and Amino-N Test Results for soils from the same county compared to Virgin Prairie | ||
| FIELD | CO2 rate ppm | Amino-N ppm |
| vegetable truck farm soil | 12.2 | 73 |
| bare, tilled tobacco soil | 27.7 | 93 |
| no-till cover-crop topsoil | 57.2 | 278 |
| no-till cover-crop subsoil | 41.5 | 110 |
| rye cover cropped soil | 45.5 | 143 |
| no-till corn soil | 47.8 | 145 |
| multi-species cover crop | 74.6 | 155 |
| virgin prairie soil, Nebraska | 78.4 | 218 |
So, nature is balancing several equations simultaneously – sequestering CO2, meeting plant needs for CO2, releasing available N and P, and dissolving soil minerals as CO2 dissolves in water and forms carbonic acid – all while furnishing soil animals with food. We should reflect on this as how modern ag science simplified it into a physical-chemical model.
Worms and roots transport OM, provide aeration and create water channels. Earthworm channels are enriched with C, nitrates and phosphates, and roots follow these channels, so under good management practices, fertility is moving downward, thanks to these interactions. In one study under organic management in Sweden, we measured biological fertility moving down at 1 inch per year. The soil was “improving itself” – this was not deep plowing or anything like that!
These long-term studies in Sweden looked at different fertility practices (control, manure, compost and three rates of conventional fertilizers) and crop rotations in the same soil type. After 17 years (the plots were kept going to their 44th year) animal manure and compost additions had tripled the respiration rate compared with control plots (with no amendments), while chemically fertilized plots had one-half to one-third the microbial respiration rate as compared even to the control plots. So fertilizers alone ultimately deprived the soil of food to the extent that the microbial population decreased and very few earthworms remained.
No-till farmers using cover crops claim that they can improve their soils better and faster than even organic, but several studies on organic farming systems that use tillage challenge this, indicating that there may be several ways to reach a goal of healthy, vibrant soils; most agree that as soon as you bring animals into the system, fertility really starts to grow.
Phosphorus Isn’t Fixed for Long!
Phosphorus is added to soils through weathering of parent material and fertilizer applications. Conventional science has taught us that this P is rapidly “fixed” in the soil and made unavailable to plants – held tightly to other elements and compounds – requiring constant P additions over time. However, while it is true that soil may complex P, it is nevertheless constantly in motion through equilibrium forces. The following figure shows factors of weathering of parent material, sorption and desorption, dissolution and precipitation, mineralization and uptake by plants, mineralization from and immobilization in humus, and leaching. What is quite surprising is that soils may contain up to 1,000 to 2,500 pounds of total P per acre; microbes may mineralize 20 to 40 pounds of P from organic pools per acre per year; and plants need only 0.3 to 0.8 ppm P – or less – in the water-soluble state at any time. Why keep fertilizing?
| Figure 2 – Dynamic Phosphorus Cycle in Agricultural Soils |
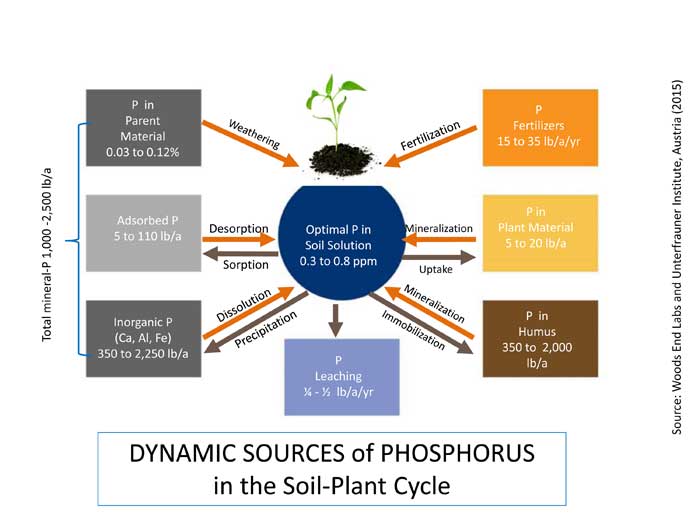 |
Recent work at Cornell by Quirine Ketterings shows that while soils do readily fix P, this capacity is limited; once soil fixation sites are saturated, due to the finite amount of soil calcium, aluminum and iron to bind it, then P may start leaching out into the soluble P pool – exceeding predictions. We’re having P pollution problems (e.g., algal blooms in the Great Lakes) perhaps because the fixing capacities of soils may be nearly saturated from decades of P use, while soil tests keep indicating more is needed.
Denmark just imposed strict P guidelines because farms there have apparently exceeded the point of P saturation. And soil tests at the University of Maine by Bruce Hoskins show that the amount of P in Maine soils is creeping up steadily because of overuse of manure and P fertilizers. Woods End is working on a test to measure biologically available phosphorus.
It Really Works – The New Science
Treating soils well in any number of ways by both organic and conventional management changes is likely to increase soil biological activity, field-level soil CO2 output, organic-N accumulation, natural nutrient cycles, and plant and animal health. And soil labs can integrate soil biology into any lab process – for example, the University of Maine Soil Testing Lab also offers a soil biology test, and Woods End has at least 25 labs nationwide now participating in offering respiration tests to improve awareness of soil biology.
The Woods End soil test integrates both nutrients and biology tests in order to arrive at a soil health score (the core idea based on five factors: respiration, amino-N, water-soluble organic carbon, aggregate stability and OM) and an overall fertility score (the health score and minimum amount of biological N available and relative P and K). Its recommendations for added fertility take into account existing available nutrients in the soil, crop needs and the likelihood of a response to added nitrogen. It also provides cover crop and lime recommendations, and Woods End does a Volumetric Aggregate Stability Test (VAST). Soils that are rich in microbes and that are not over-tilled resist falling apart in this test. Examples of soil test reports are posted at woodsend.org.
To see how your soil is doing, test soils at the same time each year. If a cover crop is in place, midsummer is a good time to test; otherwise test in late summer or early fall to see the soil in its most depleted state. Nutrient levels drift up as soils warm in spring, so testing soils too early in spring can result in excess nutrient-addition recommendations.
To illustrate nature’s brilliant system, consider that Dr. Walter Goldstein of Wisconsin has shown corn support roots dripping with exudates rich in organic compounds that, in turn, soil microbes love. This is one way that plants feed microbes. Cover crops are the magic we’ve missed in most farming practices. Nothing compares to cover cropping to improve soil, because that’s the plant communicating with your soil. Of course, Adrian Peters told us this in the 1920s, and it was overlooked.
What happens to added microbes from additions of manure and compost? The indigenous soil microflora will largely destroy the imported, exogenous microbes – meaning they are food. Comparing sources, in long-term studies, manure raised the levels of microbes a little more than compost did because compost is predigested; but compost raised the level of stable humus more. If fertilizing in the spring, choose compost for reasons including hygiene and plant safety; in late summer, use manure and let it work itself into the soil in late fall and early winter. But there’s nothing like a cover crop to do communicative soil building. Cover crops are a plant-based compost system in place, and they sequester CO2 in the soil – something you can’t do with compost.
Most studies on humus favor manure- over vegetable-based compost for long-term stability of soil carbon. Something about the protein in manure leads to a more dynamic colloid in the soil. Regarding pH, as soil OM improves, the soil system tends to regulate pH, enabling reduced limestone applications in some enhanced systems. However, plants acidify soils somewhat over time, so in some places, occasional limestone applications may help.
In summary, the overly simplified nutrient-mineral theory that has dominated agriculture for 175 years needs to be enhanced with a biological corollary that accounts for the powerful, self-regulating soil-plant community in which the soil feeds the plants. Farmers and soil labs, should they choose to do the analysis, ask for new soil testing approaches based on recognizing native mineral balances and monitoring of soil biology. Our approach to soil amending should be to foster soil life, and then we should stand back a little and let these things happen.
About the author: Will Brinton is the founder and president of Woods End Laboratories in Mount Vernon, Maine (https://woodsend.org). He spoke about soil health and soil testing at MOFGA’s 2017 Spring Growth Conference.